 Larus armenicus
Larus armenicus
(last update:
adult armenicus: May
Phenotypic characterization, taxonomic rank
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||
 Dorit Liebers & Andreas J. Helbig
Dorit Liebers & Andreas J. Helbig | Table 1: Frequency of bare part characters in adult Armenian Gulls at Lake Tuz and Lake Van. Iris: pale = unspotted pale yellowish or with up to 10% spotting. Iris: medium spotted = 10-50% spotting. Iris: dark = >50% dark spotting. |
|||
|
Tuz
|
Van
|
%
|
|
| black bill marking |
.
|
.
|
.
|
| - prominent |
182
|
259
|
95.2
|
| - faint |
?
|
19
|
4.1
|
| - lacking |
2
|
1
|
0.7
|
| leg colour |
.
|
.
|
.
|
| - intense yellow |
10
|
38
|
35.6
|
| - pale yellow |
0
|
87
|
64.4
|
| iris |
.
|
.
|
.
|
| - pale |
1
|
23
|
13.9
|
| - medium spotted |
4
|
69
|
42.2
|
| - dark |
5
|
71
|
43.9
|
Bill
Over 99% of all individuals had at least some black markings on the bill (Table 1), clearly rejecting Frede & Langbehn’s (1997) contention that armenicus lacks this character in the breeding season. However, bill markings were quite variable and were poorly developed (hard to see, small and diffuse) in about 5% of adults (n = 463). Hence, 95% of the breeding birds had a “half-moon shaped” black marking on the upper mandible (not a circle), which continued in many birds (more or less clearly) on the lower mandible. Only one single bird at Van Golu and two birds at Tuz Golu (2 out of 182) really lacked any black on the bill. The very tip of the bill, in front of the black marking, was whitish, together with the red gonys and yolk-yellow proximal part of the bill creating a four-coloured impression (Fig. 7). The red gonys spot was well developed on the lower mandible and only marginally continued on the upper mandible. The base of the bill is more intense yellow than it is in cachinnans from the Ukraine (Fig. 9), but similar to michahellis.
Leg colour
Leg colour was always yellow, but with two stages of saturation, one pale yellow (with a grey-green cast like in cachinnans) in 70% (n = 125, Table 1), the other intensely yellow (like in breeding adult michahellis) in 30% of armenicus.
Iris colour
Iris colour varied from completely pale (14%), to predominantly dark (44%) with all kinds of intermediate stages (n = 163 at Van Golu). Birds classified with a predominantly dark iris appeared “complete dark eyed” in the field at 20 m. distance (Fig. 8). At close range in the hand it appeared that a complete pale iris still showed small dark spots, and that a complete dark iris in fact was a dark spotted iris on a pale background. The orbital ring was orange-red in all birds.
Primaries
Three types of wing-tip patterns were found (Fig.18), all had black in 7 primaries and in many birds there was also black on the greater primary coverts:
* Type I with a white mirror only in P10 was most common (79%, n = 232),
* Type II with a second small white mirror on the inner web of P9 was less common (20%).
* Type III, typical of michahellis, was rare (only 2 birds, is 0.9%) in pure armenicus colonies (Fig. 19). The number of mirror spots was unrelated to sex in both armenicus and barabensis (contra Jonsson 1998). Armenian Gulls lacked a clearly delimited pale “tongue” on the inner web of P10 (the inner web was predominantly black and only at the base was a diffuse transition from this black to grey, and on P9 this tongue was much shorter and usually less clearly demarcated than in barabensis (Fig. 16, 17) or cachinnans.
Long call display
Armenian Gulls performed long call displays similar to michahellis and fuscus, never spreading the wings to an “albatross posture” typical of cachinnans. In large white-headed gulls, the long call has a stereotype pattern: it starts with one, only rarely two long elements, then miau-calls, then an intermediate call, followed by 8 to 24 short elements (Fig. 22). Long call vocalizations differed significantly from those of michahellis in 5 parameters measured sonographically (duration of first 8 short elements, frequency of miau-call and first short element, intensity distribution of harmonics and frequency modulation of miau-call), but differences were slight and gradual in nature. Several of these parameters may be related to body size, i.e. differences from michahellis may reflect smaller body size of armenicus.
Mitochondrial DNA
Phylogenetically, Armenian Gull was found to be most closely related to michahellis to the exclusion of taymirensis, barabensis and cachinnans (Fig. 24). Michahellis and armenicus are represented by two distinct clusters in the mitochondrial haplotype phylogeny, but introgression (unidirectional geneflow) was evident with two michahellis haplotypes (shaded in cluster I, Fig. 24) having invaded the armenicus population at Tuz Golu. These and a third michahellis haplotype were found in 62% of the birds in Beysehir, 14% at Tuz and 0% at Van Golu (Fig. 25). This indicates that hybridization does occur, hybrids are fertile and backcross at least with armenicus in central Anatolia. Whether geneflow in the other direction (into the michahellis population of the E Mediterranean) also occurs requires further study. Notwithstanding the ongoing introgression, the phenotypically “pure” armenicus population at Tuz and Van Golu was genetically significantly differentiated from michahellis (p < 0.0001). The genetic divergence of the two haplotype clusters (average 2% at 420 nucleotides of control domain I, Fig. 26) indicates a relatively ancient separation and long independent evolution between michahellis and armenicus, but this has so far not resulted in full reproductive isolation.
Conclusions
All evidence presented here agrees in showing that Armenian Gull does not differ qualitatively from other gulls of the argentatus-fuscus-cachinnans complex (contra Sangster et al. 1998), i.e. there is no single character allowing an unambiguous diagnosis. A combination of several characters (bill marking, wing-tip pattern, mtDNA) achieves almost 100% diagnosability in the core breeding areas of Armenian Gull (Tuz, Van), but not in the zone of secondary contact with michahellis (Beysehir), where intermediate birds occur. With respect of other taxa of the complex, in particular cachinnans, barabensis and taymirensis, diagnosability is 100% (using mtDNA and wing-tip pattern). At present it is unclear whether intrinsic barriers (mating preferences, impaired viability or fertility of hybrids) restrict the geneflow between michahellis and armenicus or whether the observed introgression is the result of recent secondary contact. The two taxa only partially fulfill the requirements of biological species status (diagnosability, reproductive isolation). However, giving the high degree of diagnosability in the core breeding areas, the limited amount of geneflow and the substantial genetic divergence between their respective mitochondrial haplotype lineages, the two taxa should be regarded as semi- or allospecies under the BSC. The situation may be comparable with the early stages of contact between argentatus and fuscus at the beginning of this century (early 1900’s), when hybridization was much more frequent than it is today. Phylogenetically, armenicus is clearly the sister taxon of michahellis (including atlantis), while it is more distantly related to barabensis, taymirensis and heuglini with which relationships have been hypothesized in the past.
Literature
Beaman, M., & S. Madge (1998): The handbook for bird identification for Europe and Western Palaearctic. London.
Buterlin, S. A. (1934): Larus taymirensis armenicus, subsp. Nov. Ibis ser. 13 (4): 171-172.
Buzun, V.A. (1993a): Armenian Gull Larus armenicus Buterlin, 1934: morpho-biometrical and behavioural distinctions with indication of taxonomical status. Russian J. Ornithol. 2: 471-490 (Russian, with English summary).
Cramp, S., & K.E.L. Simmons (1983): Handbook of the birds of Europe, the Middle East and North Africa. Bd. 3. Oxford, London, New York.
Doherty, P. (1992): Armenian Gull and Californian Gull. Birding World 5: 117-118.
Filchagov, A.V. (1993): The Armenian Gull in Armenia. British Birds 86: 550-560.
Frede, M., & H. Langbehn (1997): A contribution to the identification and distribution of the Armenian Gull. Alula 3: 102-108.
Grant, P.J. (1987): Notes on the Armenian Gull. In: International Bird Identification. Proc. 4th Intern Identification Meeting Eilat: 43.
Haffer, J. (1982): Systematik und Taxonomie der Larus argentatus-Artengruppe. In: U.N. Glutz von Blotzheim & K.M. Bauer: Handbuch der Vogel Mitteleuropas, Bd. 8. 502-515. Wiesbaden.
Hirschfeld, E. (1992): More gulls with bill bands. Birding World 5: 116.
Hume, R.A. (1983): Herring Gulls in Israel. British Birds 76: 189-191.
Jonsson, L. (1998): Yellow-legged Gulls and yellow-legged Herring Gulls in the Baltic. Alula 3: 74-100.
Kahn, N.W., J.S. John & T.W. Quinn (1998): Chromosome specific intron size differences in the avian CHD gene provide efficient method for sex identification in birds. Auk 115: 1074-1078.
Madge, S. (1990): Eine Armenienmowe Larus (cachinnans) armenicus ohne schwarze Schnabelzeichnung. Limicola 4: 216-217.
Sangster, G., C.J. Hazevoet, A.B. van den Berg & C.S. Roselaar (1998): Dutch avifaunal list: species concepts, taxonomic instability, and taxonomic changes in 1998. Dutch Birding 20: 22-32.
Satat, N., & B. Laird (1992): The Armenian Gull. Birding World 5: 32-36.
Shirihai, H., & D. Christie (1996): The Macmillan birder’s guide to European and Middle Eastern birds. London.
Snow, D.W., & C.M. Perrins (1998): The birds of the Western Palaearctic. Concise Edition. Bd. 1. Oxford, New York.
Suter, W. (1990): Comments on the breeding range of the Armenian Gull. OSME 25: 12-15.
Yesou, P., & E. Hirschfeld (1997): Which large gulls from the Larus fuscus-cachinnans-argentatus complex of (sub)species occur in Bahrain? Sandgrouse 19:111-121.
.
 Fig 1: Larus armenicus adults June 1999, Carpanak Island, Van Golu, Turkey. Picture: Andreas J. Helbig.
Fig 1: Larus armenicus adults June 1999, Carpanak Island, Van Golu, Turkey. Picture: Andreas J. Helbig.  Fig 2: Larus armenicus breeding island in northern Tuz Golu, central Anatolia, Turkey, May 1999. Picture: Andreas J. Helbig.
Fig 2: Larus armenicus breeding island in northern Tuz Golu, central Anatolia, Turkey, May 1999. Picture: Andreas J. Helbig. Fig 3: Larus armenicus mixed with michahellis breeding island in Beysehir Golu, Turkey, May 1999. Picture: Andreas J. Helbig.
Fig 3: Larus armenicus mixed with michahellis breeding island in Beysehir Golu, Turkey, May 1999. Picture: Andreas J. Helbig. Fig 4: Larus armenicus at Carpanak Island, Van Golu. Largest breeding colony in Turkey holding 2.000 pairs, June 1999. Picture: Andreas J. Helbig.
Fig 4: Larus armenicus at Carpanak Island, Van Golu. Largest breeding colony in Turkey holding 2.000 pairs, June 1999. Picture: Andreas J. Helbig. Fig 5: Larus armenicus adults June 1999, Carpanak Island, Van Golu, Turkey. Picture: Andreas J. Helbig. Details on methods and material.
Fig 5: Larus armenicus adults June 1999, Carpanak Island, Van Golu, Turkey. Picture: Andreas J. Helbig. Details on methods and material. Fig 6: Larus armenicus adult June 1999, Carpanak Island, Van Golu, Turkey. Picture: Andreas J. Helbig.
Fig 6: Larus armenicus adult June 1999, Carpanak Island, Van Golu, Turkey. Picture: Andreas J. Helbig. Fig 7: Larus armenicus female at Tuz Golu, Turkey, May 1999. Picture: Andreas J. Helbig.
Fig 7: Larus armenicus female at Tuz Golu, Turkey, May 1999. Picture: Andreas J. Helbig. Fig 8: Larus armenicus male at Tuz Golu, Turkey, May 1999. Picture: Andreas J. Helbig.
Fig 8: Larus armenicus male at Tuz Golu, Turkey, May 1999. Picture: Andreas J. Helbig. Fig 9: Larus cachinnans female at Asow Sea, Ukraine, April 1997. Picture: Dorit Liebers.
Fig 9: Larus cachinnans female at Asow Sea, Ukraine, April 1997. Picture: Dorit Liebers. Fig 10: Larus (ssp?) barabensis male at Tengiz Lake, Kazachstan, May 1998. Picture: Andreas J. Helbig.
Fig 10: Larus (ssp?) barabensis male at Tengiz Lake, Kazachstan, May 1998. Picture: Andreas J. Helbig. Fig 11: Larus armenicus adult June 1999, Carpanak Island, Van Golu, Turkey. Picture: Andreas J. Helbig.
Fig 11: Larus armenicus adult June 1999, Carpanak Island, Van Golu, Turkey. Picture: Andreas J. Helbig. Fig 12: Larus armenicus adult June 1999, Carpanak Island, Van Golu, Turkey. Picture: Andreas J. Helbig.
Fig 12: Larus armenicus adult June 1999, Carpanak Island, Van Golu, Turkey. Picture: Andreas J. Helbig. Fig 13: Larus armenicus adult June 1999, Carpanak Island, Van Golu, Turkey. Picture: Andreas J. Helbig.
Fig 13: Larus armenicus adult June 1999, Carpanak Island, Van Golu, Turkey. Picture: Andreas J. Helbig. Fig 14: Larus armenicus male at Tuz Golu, Turkey, May 1999. Picture: Andreas J. Helbig.
Fig 14: Larus armenicus male at Tuz Golu, Turkey, May 1999. Picture: Andreas J. Helbig. Fig 15: Larus armenicus female at Tuz Golu, Turkey, May 1999. Picture: Andreas J. Helbig.
Fig 15: Larus armenicus female at Tuz Golu, Turkey, May 1999. Picture: Andreas J. Helbig. Fig 16: Larus (ssp?) barabensis male at Tengiz Lake, Kazachstan, May 1998. Picture: Andreas J. Helbig.
Fig 16: Larus (ssp?) barabensis male at Tengiz Lake, Kazachstan, May 1998. Picture: Andreas J. Helbig. Fig 17: Larus (ssp?) barabensis female at Tengiz Lake, Kazachstan, May 1998. Picture: Andreas J. Helbig.
Fig 17: Larus (ssp?) barabensis female at Tengiz Lake, Kazachstan, May 1998. Picture: Andreas J. Helbig. Fig 18: Larus armenicus three types of wing-tip patterns. Illustrations: Dorit Liebers.
Fig 18: Larus armenicus three types of wing-tip patterns. Illustrations: Dorit Liebers.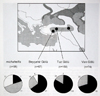 Fig 19: Larus armenicus and michahellis wing-tip pattern distribution.
Fig 19: Larus armenicus and michahellis wing-tip pattern distribution. Fig 20: Larus armenicus at Tuz Golu, Turkey, May 1999. Picture: Andreas J. Helbig.
Fig 20: Larus armenicus at Tuz Golu, Turkey, May 1999. Picture: Andreas J. Helbig. Fig 21 Head+Bill length plotted against Wing length for three taxa.
Fig 21 Head+Bill length plotted against Wing length for three taxa.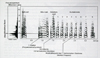 Fig 22: Larus armenicus long call.
Fig 22: Larus armenicus long call. Fig 23: Miau-calls within the long calls of armenicus and michahellis.
Fig 23: Miau-calls within the long calls of armenicus and michahellis. Fig 24: Reconstruction of geneology tree for armenicus and michahellis.
Fig 24: Reconstruction of geneology tree for armenicus and michahellis.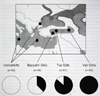 Fig 25: Presence of mitochondrial haplotytpes in Turkey and in the E. Mediterranean.
Fig 25: Presence of mitochondrial haplotytpes in Turkey and in the E. Mediterranean. Fig 26: Number of nucleotide differences.
Fig 26: Number of nucleotide differences.